26 Dec 2016
Serotonin é o biogenic amine que se forma de tryptophan amino ácido pelo seu hydroxylation e decarboxylation. Uma quantidade significativa de um serotonin contém nas células enterokhromaffinnykh do intestino, CNS, principalmente em hypothalamus e mesencephalon, trombócitos, mais pequena quantidade no labrotsitakh, células de mastro, glândulas supra-renais. Um serotonin exerce o impacto em atividade nervosa, redução de causas de um musculation liso de um intestino, útero, brônquios, e também restrição de navios. As reações de um organismo a um serotonin são a pedra angular o central, myotropic, ganglionic, efeitos reflexivos.
Educação. Um serotonin (5-gidroksi-triptamin, 5-HT) sintetiza-se nas células de epitélio internas enterokhromaffinnykh de um L-tryptophan. Um serotonin também forma-se em células nervosas de mezenterialny plexus e em um CNS onde desempenha um papel de mediador. Os trombócitos não sintetizam um serotonin, contudo tomam-no e acumulam-se.
Receptores de Serotonin. Há vários tipos de receptores. Dos receptores de ponto de vista farmacológicos 5-HT1 e 5-NT2, e também 5-HT3 e 5-HT4 são importantes. Por via de regra, os receptores funcionam pela G-proteína. O receptor 5-HT3 representa o canal cationic.
Efeito de serotonin. Sistema cardiovascular. A influência de serotonin no sistema cardiovascular é complexa como o serotonin tem o efeito (às vezes em frente enviado) em muitos sítios por tipos diferentes de receptores. Por 5-HT2-retseptory localizado em músculos não estriados de paredes de navios, um serotonin causa a restrição de navios. O efeito ao mesmo tempo indireto de serotonin pode levar a vasodilatation e a depressão do ABP: por 5-HT1A-retseptory bloqueia neurônios compreensivos de um brainstem e a periferia que leva à depressão de um tonus compreensivo; a ativação de receptores 5-HT1 em um endothelium vascular leva à emissão de vazodilatator (prostatsickin, NÃO). O serotonin que se lança de trombócitos desempenha um papel em processos de thrombogenesis, hemostasis e em hypertonia pathogenesis em mulheres grávidas.
Sumatriptan (um agente contra a enxaqueca) é um 5-HT1D agonist e 5-HT1B-retseptorov. A restrição de navios craniais causados pelo bloqueio da emissão de neuropeptids leva a um assim chamado "neurogenic inflamação" ou dirigir a crise vascular. Ao mesmo tempo pode haver uma sensação do constrangimento em um peito atado a uma censura espasmódica de artérias coronárias. Outros "tryptones" são naratriptan, zolmitriptan e rizatripan.
Tratado digestivo. O serotonin formado em mezenterialny plexus ou células enterokhromaffinnykh estimula um peristalsis e a substância segreda de um intestino por meio da influência em 5-HT4-retseptory.
As tentativas do impacto farmacológico em um motility de um intestino por receptores serotonin foram ineficazes por enquanto. Tsizaprid, 5-HT4 agonist, contudo, pode aumentar motility propulsivo de um intestino. Este efeito de lado bloqueia-se por Atropinum que demonstra a insurreição da concentração de Acetylcholinum abaixo da influência de tsizaprid. A droga dá o grupo de efeitos de lado. É inactivated no mecanismo com a participação de SURZA4 que leva a várias interações imprevistas com outras substâncias. Tsizaprid causa perturbações (inclusive o sério) um ritmo cordial (a redução de intervalos QT), por isso, a droga não encontrou a aplicação.
Sistema nervoso central. Os receptores de Serotonin desempenham um papel importante em várias funções CNS.
Fluoxetine é o antidepressivo que bloqueia o regresso serotonin recompreensão. Consideravelmente aumenta a motivação, e também abaixa o apetite.
Sibutramin, blocker do regresso recompreensão serotonin neuronial e Noradrenalinum; aplica-se como remédio pela obesidade.
Ondansetron tem o efeito antiemético expresso no vômito causado pela aplicação de tsitostatik. É antagonista de 5-HT3-retseptorov. Os análogos de ondansetron são tropisetron e granisetron.
O ácido e os outros de psychedelia (psikhotomimetika), como mescalina e psilocybin, causam alucinações, desordens de consciência, medo, possivelmente, por causa da ativação de 5-NT-retseptorov.
Efeitos de serotonin
O pessoal de universidade de McMaster tornou-se autores do trabalho científico durante o qual o gene-chave da obesidade - uma espécie de serotonin se encontrou. É bem conhecido que serotonin é "o hormônio de alegria", o seu desenvolvimento por um cérebro fomenta a estabilidade emocional e o bom humor. Mas, como os pesquisadores canadenses explicam, o serotonin que é responsável por emoções agradáveis pertence ao primeiro tipo desta conexão.
"Divide-se em dois tipos: na cena de ação e em uma forma de síntese. O primeiro tipo desenvolve-se em um cérebro e influi em emoções diferentes", - os biólogos explicaram.
Ao segundo tipo, serotonin periférico pertence esta substância regula a atividade do tecido gorduroso marrom do qual o desenvolvimento da obesidade depende. Você também pode gostar de Pankramin.
A gordura marrom contém componentes que fomentam a redução no açúcar no sangue, e também à ardência de calorias e o seu processamento na energia. Em um corpo da pessoa há certas zonas onde a gordura marrom se instala e as suas jaulas são mais ativas, o número na pessoa é mais harmonioso. Os pesquisadores canadenses vieram à conclusão que suprimindo serotonin do segundo tipo; é possível aumentar a atividade consideravelmente metabólica de células do tecido gorduroso marrom. À sua vez, forçará um corpo "a queimar" a gordura branca � e, ocorrerá independente de como a comida de alta caloria se come pela pessoa.
Receptores de Serotonin
Os efeitos de serotonin são extremamente vários. Esta substância serve de um mediador em TsNS, os músculos lisos de influências dos navios e um GIT, participa em trombotsitarnom vascular um homeostasis. Os métodos da clonagem molecular revelaram o inesperadamente grande número os receptores serotonin que baseado em estrutura e função podem dividir-se em 4 tipos. 5-T1--, 5-HT2-and 5-HT4-retseptory integra-se à G-proteína e por esta proteína e os sistemas correspondentes dos segundos intermediários influem em funções de várias enzimas e propriedades elek-trofiziologich de jaulas effector. Ao contrário, 5-HT3-retseptory atam-se com canais de íon. Aqui consideraremos stimulators e blockers os receptores serotonin. A medicina mais nova destes grupos que fazem funcionar seletivamente em subtipos separados os receptores serotonin recebeu-se em trabalhos com o uso de receptores recombinant. Também pararemos em modelos experimentais que aplicam a uma pesquisa dos meios que influem em funções mentais difíceis e a sua persistência de violações, comportamento agressivo, preocupação, depressão, vigilância de sonho de ciclo e outro. stimulators seletivos modernos de subtipos separados os receptores serotonin já com o êxito se aplicam em caso de enxaqueca e preocupação e o blockers seletivo em caso de um número de violações gastrintestinais. É possível influir em efeitos fisiológicos de serotonin também por meio dos meios que produzem transferência de serotoninergich. Deste modo, os nervos inibidores da captura de regresso de serotonin foram a medicina eficaz do tratamento de depressão e preocupação.
Apesar de que o papel serotonin em muitos processos fisiológicos e patológicos não levante dúvidas, os pontos da sua aplicação e os mecanismos da ação estudam-se mal. Possivelmente, tal situação se causa em parte pela variedade o serotoninovykh de receptores. Estes receptores gravados no começo por métodos farmacológicos recebem-se por KDNK que se clona hoje. Os receptores de Recombinant serotoninovy usam para estudar de mecanismos moleculares do efeito de serotonin, e também para a pesquisa dos agentes que influem seletivamente em subtipos separados destes receptores. O círculo de um uso clínico de agentes semelhantes fica mais largo e mais largo.
Informação histórica
Em 1930o Erspamer começou a estudar a localização de células enterokhromaffin por meio de manchas em derivados de índole. A concentração mais alta de tais derivados gravou-se no mucosa por um GIT; além disso houve trombócitos e alguns departamentos de um CNS (Erspamer, 1966). Daqui a pouco Peydzh e os co-autores, que trabalham na clínica de Clevelend, pela primeira vez emitiram a substância vasoconstrictive lançada de trombócitos no decorrer de uma paragem de hemorragia e decifraram a sua estrutura (Concordância et al., 1948). Esta substância denominada por Paige um serotonin (Page, 1976), apareceu àqueles derivados de uma índole que se investigou por Erspamer. A descrição de caminhos da síntese e desintegração de um serotonin (Uden-amigo, 1959) e as suas propriedades angiotonic (Sjoerdsma, 1959) permitiu fazer uma hipótese segundo a qual as implicações de uma assim chamada síndrome carcinoid em pacientes com tumores de células enterokhromaffin se causam pela produção aumentada desta substância. Realmente, tais pacientes têm uma ejeção diária com a urina de um serotonin e o seu metabolites pode conseguir centenas de miligramas. Alguns sintomas desta doença até certo ponto indicam mecanismos do efeito de serotonin. Deste modo, em pacientes a psicose semelhante ao ácido que surge na recepção pode desenvolver-se. Considerando que em animal e substâncias de tecidos vegetais com a ação de alucinógeno se encontram semelhantes a tripta-miny, é possível supor que as substâncias semelhantes se formem e causem uma sintomatologia psicótica em pacientes com a síndrome characinoid. Sobre a função de mediador de um serotonin em um cérebro de mamíferos sugeriu-se a meados de anos 50 (Brodie e Shore, 1957).
Os primeiros dados sobre mecanismos moleculares do efeito de serotonin obtiveram-se em experiências em um fígado de Fasciola hepatica (Mansour, 1979). Abaixo da influência de serotonin nele a mobilidade e a concentração de tsAMF agudamente aumentaram; tanto isto, como o outro efetua bloquearam-se por um ácido. O aumento na mobilidade causou-se por phosphorylation tsAMF-dependente de fosfofruktokinaza que limita glycolysis enzima. Contudo os receptores serotoninovy que medeiam estes efeitos em um fígado, provavelmente, outros, do que receptores de mamíferos inter-relacionados com adenilattsiklaz a serotonin. No máximo assim dados detalhados em mecanismos do efeito de serotonin não se conseguiu para receber por enquanto.
Serotonin apareceu em plantas e animais já em primeiras etapas da evolução, e a estes, possivelmente, a abundância de receptores serotonin fala (Peroutka e Howell, 1994). A clonagem destes receptores mostrou que alguma medicina que se considerou antes seletiva em relação aos seus subtipos separados de fato tem a alta afinidade em relação a vários subtipos. Mais detalhadamente sobre história de estudar e efeitos de serotonin ver o artigo Sjoerdsma e Palfreyman (1990).
Propriedades químicas de serotonin
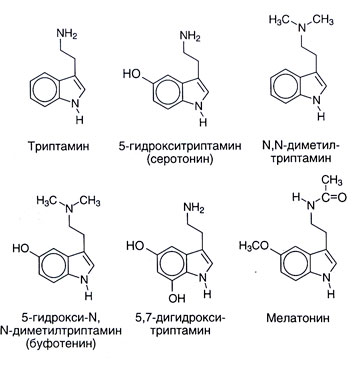
Fontes. A estrutura química de serotonin e algumas conexões fechadas a ele dá-se no figo. 11.1. Serotonin é comum em uma vida de plantas e animais: encontra-se em vertebrado, tunicates, moluscos, arthropods, celenterados, no fruto e em nozes. Também encontra-se em venenos � em uma urtiga, em vespas e escorpiões. Os parentes sintéticos ou naturais numerosos à substância serotonin também em um grau variado têm os efeitos fisiológicos centrais e periféricos. Muitos Nem O-metilirovannye indolamines (por exemplo, N, N-dimetiltriptamin) são alucinógenos. Como podem desenvolver-se em um organismo, muito tempo considerou-os possíveis responsável por pelo menos algumas exposições de psicoses. Melatonin (5 metoksi N atsetiltriptamin) forma-se de serotonin por N-acetylation com o O-methylation subsequente (figo. 11.2). Esta substância serve do indolamine principal do corpo onde a sua síntese se regula por fatores externos (especialmente, nível de iluminação). Melatonin causa depigmentation de melanotsit da pele e suprime a função de ovários. Possivelmente, desempenha um papel em biorritmos e por isso pode ser útil em uma síndrome da modificação de fusos horários.
Síntese e catabolism. Um serotonin forma-se do ácido amino insubstituível de um tryptophan em 2 etapas (figo. 11.2). Na primeira etapa abaixo da influência de triptofangidroksilaza 5 hydroxytryptophan formam-se, é a limitação serotonin reação de síntese. Triptofangidroksilaza representa um oxidase com as funções mistas. O oxigênio molecular, e como coenzima tetrahydrobiopterine toma parte na reação catalisada por ele. A atividade de triptofangidroksilaza, bem como tyrosinehydroxylase, regula-se por phosphorylation, contudo o triptofangidroksilaza não se inibe pelo produto final no mecanismo do comentário negativo. Em um cérebro de triptofangidroksilaz não se satura com substrate e por isso a tarifa da síntese de um serotonin depende da concentração de um tryptophan. O último vem a células cerebrais pela captura ativa por meio da transportadora que é responsável pela transportação de vários ácidos amino neutrais e ramificados. Neste sentido a manutenção de um tryptophan em um cérebro depende não só da sua concentração no plasma, mas também na concentração de outros ácidos amino que competem com um tryptophan por uma transportadora.
Decarboxylation leva 5 hydroxytryptophans à formação de um serotonin. A disputa longa em se decarboxylases 5 hydroxytryptophans e DOFA são diferentes ou a mesma enzima, resolveu-se por KDNK métodos se clonam resultou que o mesmo produto genético é responsável por decarboxylation de ambos substrates. Agora esta enzima chama-se decarboxylase de ácidos L-amino aromáticos. É extremamente comum e afeta muitos substrates. 5 hydroxytryptophan são decarboxylized muito rapidamente e em um cérebro não se encontra quase. Neste sentido as tentativas de afetar a concentração em um cérebro serotonin pela concentração modificam-se 5 hydroxytryptophans condenam-se ao fracasso.
O caminho principal de catabolism serotonin a transformação no ácido 5-hydroxyindolacetic que também prossegue em 2 etapas (figo. 11.2). No início abaixo da influência de MAO o 5-hydro-xyindolacetaldehyde que então passa no ácido 5-gidroksi-indolacetic abaixo da influência da enzima aldegiddegidrogenaza comum em um organismo forma-se (a quantidade insignificante 5 hydroxyindolacetaldehydes transformam-se no álcool � 5-gidroksitriptofol). O ácido 5-hydroxyindolacetic é activly retirado de um cérebro; este processo suprime-se com um nervo inibidor não-específico de probenetsidy de transferência de transepithelial. Como em células nervosas quase 100% de todo o metabolites de uma queda de serotonin à ação de ácido 5-hydroxyindolacetic, a tarifa de um circuito de um serotonin em um cérebro prevê-se na insurreição do nível de ácido 5-hydroxyindolacetic depois da introdução de probenetsid. O ácido 5-hydroxyindolacetic que se forma em um cérebro e outros órgãos, e também as poucas quantidades do 5-gidroksitriptofol e glucuronides se retira com a urina. Normal o diário egestion do ácido 5-hydroxyindolacetic no adulto faz 2 � 10 mgs. Valores mais altos � um sinal fiável de síndrome carcinoid. A síntese agudamente aumentada de um serotonin nesta doença exige grandes números de pyridinic nucleotides e um tryptophan e por isso sinais da deficiência de ácido nicotinic e um tryptophan não uma raridade em tais pacientes. A insurreição de causas de etanol da manutenção de NADN, e em consequência de 5 hydroxyindolacetaldehyde passa de um caminho que se oxida de um catabolism à recuperação (figo. 11.2).
Estes são levanta egestion de um 5-gidroksitriptofol um pouco e respectivamente reduz egestion de ácido 5-hydroxyindolacetic.
Há dois isoenzymes de MAO: MAO A e MAO B. No início dividiram-se baseados em afinidade a substratos e sensibilidade a nervos inibidores; agora tanto os isoenzymes clonam-se, como as propriedades das formas clonadas e naturais foram idênticas (Shih, 1991; também ver hl. 10). MAO A tem a afinidade preferencial a serotonin e noradrenaline, e o seu nervo inibidor seletivo é clorgilin. MAO B mais trabalhos em β-fenil-etilamin e benzylamine; MAO B nervo inibidor seletivo selegilin. A afinidade tanto de isoenzymes a dopamine como de triptamin é idêntica. As jaulas nervosas contêm tanto MAO A como MAO B � principalmente em uma membrana externa de mitochondrions. Como o isoenzyme principal de plaquetas que também contêm serotonin na alta concentração serve MAO B.
Supôs-se que também há outros caminhos de um catabolism de serotonin, por exemplo sulphation e sobre - ou N-methylation. A viagem final, especialmente, pode levar à formação de uma substância psychotropic endógena 5-hydroxy-N, N-dimetiltriptamina (bufotenina, figo. 11.1). Contudo outros metilirovanny indolamines (N, N-dimetiltriptamin, 5-metoksi-N, N-dimetiltriptamin) a maior parte mais exprimiram propriedades de alucinógeno, e o seu papel em pathogenesis de psicoses é mais provável.
O inactivation de serotonin executa-se não só pela desintegração enzymatic, mas também por meio da captura de regresso. Na + - a transportadora dependente localizada em uma superfície externa da membrana presynaptic da terminação serotoninergich (fornece a remoção de serotonin de uma fenda sinótica) e uma superfície externa de uma membrana de plaquetas é responsável por esta captura (extrai serotonin do sangue). Em plaquetas é o caminho único da ação de reabastecimento de serotonin como não há enzimas da síntese desta substância neles. A transportadora serotonin, bem como outras transportadoras de monoamines, clona-se (hl. 12).
Pontos de aplicação serotonin
A maior parte de parte de toda a manutenção de serotonin em um organismo cai à ação de tecidos periféricos embora também sirva como um mediador em um CNS. A sua concentração em células enterokhromaffinnykh e trombócitos é a mais alta. Um serotonin desempenha um papel importante em uma regulação de um motility de um GIT.
Células de Enterokhromaffinny. Estas células sossegam no mucosa um GIT. Especialmente há muitos deles em um duodeno. Nas células enterokhromaffinnykh sintetiza-se de um tryptophan e um serotonin reúne-se, e também outros agentes biologicamente ativos, por exemplo a substância P e kinina contêm. Há certo nível da substância segreda de base de um serotonin em um GIT. Esta substância segreda amplifica no momento do estiramento mecânico (por exemplo, entrando em uma nutrição ou solução de sal hipertônica) e em uma maçante de fibras de motivo de nervos vagus. Possivelmente, o efeito estimulante de serotonin em um motility de um GIT também se medeia pela sua influência em neurônios de um plexus intermuscular (Gershon, 1991; também ver hl. 38). A substância segreda agudamente aumentada de um serotonin e outros agentes biologicamente ativos na síndrome carcinoid segue-se das perturbações gastrintestinais, cardiovasculares e nervosas correspondentes. Além disso, a síntese aumentada de um serotonin pode resultar na deficiência de ácido nicotinic e um tryptophan.
Trombócitos. Os trombócitos diferenciam-se de outros elementos formulados de um sangue, especialmente, na capacidade de tomar a loja e lançar um serotonin. A síntese de um serotonin em trombócitos não acontece. Um serotonin é trombócitos tomados de um sangue e vem para o armazenamento ao segredo elektronop grânulos por meio do transporte ativo. Estes processos são em muitos aspectos semelhantes a captura e armazenamento de Noradrenalinum nas terminações compreensivas (hl. 6 e 12). Por uma membrana de trombócitos um serotonin transfere-se por meio de Na + - transporte dependente, e a grânulos � pelo transporte ativo secundário com o uso como uma fonte de energia de um declive eletroquímico de H + enquadrado por N +-Atfazoy. Ao mesmo tempo a concentração de um serotonin em grânulos consegue 0.6 mol/l, é 1000 vezes mais alto, do que em cytoplasm de trombócitos. Tarifa em + - recompreensão serotonin dependente por trombócitos um indicador sensível de atividade de nervos inibidores de recompreensão serotonin.
A função principal de trombócitos é hemostasis: fecham fendas no endothelium danificado. De outro lado, a integridade de um endothelium desempenha um papel importante no funcionamento de trombócitos (Furchgott e Vanhoutte, 1989). O endothelium constantemente contata a trombócitos como por causa das forças de turno que agem no sangue atual deslocam-se à periferia de navios (Gibbons e Dzau, 1994). O efeito de Vasoconstrictive de serotonin e tromboksan de A2 resistem-se por um fator endothelial de vasorelaxation (N0 e, possivelmente, algumas outras substâncias) (Furchgott e Vanhoutte, 1989; figo. 11.4). Para adesão e agregação de trombócitos a condição de um endothelium tem a importância crucial (Hawiger, 1992; Ware e Heistad, L993). Quando os trombócitos estão contíguo ao endothelium danificado, emitem as substâncias que causam a sua adesão e o lançamento de um serotonin. ADF e ò ð îìáîêñàí A2 pertencem a tais substâncias (hl. 26 e 55). Linkng de um serotonin com 5-HT2A-retseptorami tem o efeito proagregant débil que amplifica agudamente na presença de um collagen. Se o defeito de uma parede vascular conseguir camadas de músculo lisas, então um serotonin dá o efeito vasoconstrictive direto que serve de um de mecanismos hemostasis. Este efeito amplifica abaixo da influência de agentes biologicamente ativos que se emitem no campo do dano tromboksan de A2, kinin, vasoactive péptidos. A formação de thrombi em uma aterosclerose fomenta-se pela destruição de um endothelium e, por conseguinte, a falta de um fator endothelial de vasorelaxation. Nestas condições a condução de processos a thrombogenesis prosseguem incontroladamente, como um círculo vicioso. Um papel neles também desempenha-se por um serotonin. O quadro semelhante pode observar-se em outras doenças de navios, por exemplo a síndrome de Reynaud e vasospastic stenocardia.
A descrição a figo. 11.4. Funções de um serotonin de trombócitos. O lançamento de um serotonin de trombócitos começa-se eles por adesão e agregação. À sua vez, um serotonin causa 1) a ativação trombócitos de á-ÍÒ-receptors e, vrezultata, a modificação de uma forma e aceleração da agregação do último, 2) ativação 5-HT, - os receptores semelhantes do endothelium com a alocação de um fator endothelial de vasorelaxation, 3) a ativação de S-HT-receptors de músculos não estriados de navios e restrição do último. Todos estes processos procedem na interação com muitos outros agentes biologicamente ativos e consequentemente levam à hemorragia de paragem.
Sistema cardiovascular. A reação típica de vasos sanguíneos a serotonin estreita-se. Os navios de órgãos de GIT, rins, pulmões e cérebro são especialmente sensíveis a ele. Serotonin também causa a redução de músculos não estriados de brônquios. Os seus efeitos sobre o coração são vários que a modificação de receptores serotonin de tonus de nervos vegetativos e reações reflexivas se explica pela ativação de subtipos diferentes (Saxena e Villalon, 1990). Deste modo, chronotropic positivo direto e o efeito inotropic de serotonin no coração podem disfarçar-se por efeitos da exaltação das fibras que vão de receptores de barra e chemoceptors. A influência de um serotonin nas terminações afferent de nervos vagus causa Yarisha reflexivo de Betsold que se mostra um bradycardia agudo e a queda do ABP. Às vezes os arterioles abaixo da influência de um serotonin não se estreitam, e, ao contrário, a supressão do lançamento de Noradrenalinum das terminações compreensivas estendem-se em consequência da alocação de um fator endothelial de vasorelaxation e Prostaglandinums, e também. De outro lado, um serotonin em si mesmo fortalece a ação vasoconstrictive de Noradrenalinum, angiotensin 11 e Histaminum. Fomenta o efeito estático haemo até mais eficaz de serotonin (Gershon, 1991).
GIT. Provavelmente, como a fonte principal e o armazenamento de um serotonin em um organismo servem células enterokhromaffinny mucosas um GIT. O serotonin alocado por estas células consegue uma veia porta a um fígado onde se metaboliza abaixo da influência de MAO A (Gillis, 1985). Algum montante de serotonin passa um metabolismo hepatic, mas toma-se rapidamente um endothelium de tubos capilares pulmonares e também se afeta por MAO. O serotonin que se aloca em uma parede de órgãos de um GIT no momento do seu estiramento mecânico ou a exaltação de nervos vagus participa em uma regulação local destes órgãos. Abaixo da influência de um serotonin um motility de um estômago e um intestino pode amplificar tanto, e frear-se (Dhasmana et al., 1993) como em um GIT há pelo menos 6 subtipos os receptores serotonin (etiqueta. 11.2). O efeito estimulante de serotonin causa-se pela sua ação nas terminações dos nervos convenientes a camadas musculares longitudinais e circulares (5-HT4-retseptory), em neurônios entre muros (5-HTj-and 5-NT|R-retseptory) e é direto em músculos lisos (5 receptores da IO em intestinos e 5-HT2B-retseptory no dia de um estômago). Em uma garganta o serotonin afeta 5-HT4-retseptory que em tipos diferentes de animais pode seguir-se tanto reduzindo, como o relaxamento de músculos lisos. Os 5-HT3-retseptory (são muitos visitantes nas terminações de fibras sensíveis da vagabundagem e outros nervos, e também nas jaulas) desempenham um papel-chave em um reflexo emético (Grunberg e Hesketh, 1993). Em uma textura intermuscular as terminações serotoninergetic encontram-se. O lançamento de serotonin em intestinos causa-se por acetylcholine, a irritação de nervos compreensivos, e aumento na pressão intestinal intra e redução no pH (Gershon, 1991). O serotonin que se emite ao mesmo tempo, à sua vez, começa a redução peristáltica.
A área principal da concentração de corpos de neurônios serotonin em CNS é os núcleos da costura de brainstem. Os processos destes neurônios vão a todos os departamentos de uma corda dianteira e espinal (hl. 12). Um serotonin aloca-se não só nas terminações presynaptic, mas também em um assim chamado varicosity de axons onde não há exatamente synapses expressos (Descarries et al., 1990). Nestes casos afeta muitas estruturas adjacentes ao mesmo tempo. Tal característica da alocação e o efeito de serotonin vão se compor com o ponto de vista comum que um serotonin não é só um mediador, mas também o modulador de neurônio (hl. 12).
Nas terminações de neurônios serotonin há todos componentes necessários para a síntese de um serotonin de um tryptophan (figo. 11.2). O serotonin formado rapidamente vem a bolhas sinóticas onde nele MAO não pode trabalhar. Depois do lançamento em uma racha sinótica um serotonin toma-se novamente a terminação nervosa por meio de Na - uma transportadora dependente. Esta captura de regresso serve de um caminho eficaz de um inactivation de um mediador. As mesmas moléculas de um serotonin que não voltam à terminação nervosa arruinam-se por MAO localizado em neurônios postsynaptic e as seguintes células.
Efeitos eletrofisiológicos. Estes efeitos de um serotonin diferenciam-se em áreas diferentes de um cérebro e em neurônios diferentes e dependem em que receptores afeta (etiqueta. 11.3; Aghajanian, 1995). Um serotonin pode ter a excitação e efeito de freio que dissente na dinâmica temporária nos mesmos neurônios. Deste modo, em neurônios hippocampus um serotonin causa no início a polarização hyper (causado pela ativação de 1A-receptores 5-HT), então a depolarização lenta (causado pela ativação de 5-HT4-retseptorov).
TsNS. Serotonin influi em TsNS muitas funções, inclusive um sonho, atividade cognitiva, percepção, gestão de movimentos, controle termal, sensibilidade dolorosa, apetite, comportamento sexual e regulação endócrina. Em um cérebro todos clonaram-se os receptores serotonin encontram-se, e muitas vezes no mesmo departamento há vários receptores. Além disso, embora a expressão de receptores serotonin em neurônios separados se estude insuficientemente, é possível acreditar que no mesmo neurônio vários subtipos destes receptores podem localizar-se, e a sua ativação pode seguir-se tanto de synergistic como de efeitos antagônicos. Pode ser a causa de uma variedade extraordinária de influências de serotonin em funções cerebrais.
Polarização de Hyper e redução na resistência de uma membrana que surge em caso da ativação 5-HT) A-receptores, causam-se pelo aumento na permeabilidade de potássio. Estes efeitos bloqueiam-se pela toxina, mas não dependem de tsAMF. Por isso, podem causar-se pela interface direta de 5-NT|A-retseptorov (pela G-proteína semelhante a Gi) com o canal de potássio (Andrade et al., 1986). A ativação dos receptores localizados em um corpo e as dendrites dos neurônios de núcleos de uma costura de um tronco de um cérebro também leva a K + - polarização hyper dependente. Aqui também participa em uma transmissão j de um sinal de um receptor ao canal sensível à toxina de G-proteína, mas a corrente de potássio tem outras características, do que o que surge em caso da ativação de postsynaptic 5-HT1A-retseptorov em um hippocampus. Ainda não se conhece por meio de que ativação de mecanismos 5-HT1D-ayTopeuerrropoB leva à supressão de lançamento de serotonin. Provavelmente, o número dos canais calcic que se abrem em resposta a impulso nervoso as reduções potenciais.
A ativação de 5-HT2A-retseptorov segue-se da depolarização lenta. Em alguns departamentos de um cérebro (por exemplo, no córtex prefrontal, um núcleo adjacente e um núcleo de motivo de um nervo facial) esta depolarização causa-se pela redução na permeabilidade de potássio (Aghajanian et al., 1987). Também há outro mecanismo unido com a influência em canais de íon; leva ao crescimento de excitabilidade do neurônio e fortificação da reação a mediadores excitantes (por exemplo, um glutamate). O papel do sistema fosfoinozitid nestes efetua ainda não se estabelece. Quando no mesmo neurônio também há 5-HT1-retseptory e 5-Nt2d-retseptory, a reação final a serotonin depende de uma proporção entre a polarização hyper causada pela ativação de 5-HT1-retseptorov e a depolarização causada pela ativação de 5-HT2A-retseptorov. No contexto da polarização 5-Nt2d-blokatorov hyper amplifica. Em muitas áreas do latido 5-HT2A-retseptory localizam-se em neurônios inseridos de Gamkergichesky e em neurônios piramidais. Por isso, a estimulação destes receptores pode levar a influências multidirecionais em neurônios piramidais na dependência porque se a ação nestes neurônios ou em neurônios de Gamkergichesky prevalecerá. A ativação de 5-Nt2s-retseptorov em oocytes da rã, rápido do m do ARN destes receptores, leva à supressão da corrente de potássio. Em um cérebro tal efeito ainda não se revela. A estimulação de 5-HT4-retseptorov que se segue da ativação também causa a depolarização lenta causada pela redução na permeabilidade de potássio em neurônios. Ainda não é claro porque dois tipos diferentes de receptores serotonin inter-relacionados a vários sistemas dos segundos intermediários dão o mesmo efeito fisiológico. Além disso, a depolarização lenta causa-se pela ativação de um mais tipo de receptores serotonin 5 HT1P-receptores. Estes receptores acompanham-se pelo disponível só em neurônios intestinais intra e têm propriedades farmacológicas especiais (Gershon, 1991).
A ativação de 5-HT3-retseptorov causa a depolarização bystry. Causa-se abrindo-se do canal que passa Na + e K + (Higashi e Nishi, 1982) e faz um complexo uniforme (hemo canal sensível) com um receptor. A organização semelhante de um semelhante 5-HT3-retseptora à organização N-holinoretseptora confirmou-se em experiências com a fixação local. Os 5-HT3-retseptory encontram-se em TsNS, gânglios compreensivo, fibras afferent paracompreensivas e compreensivas, intra neurônios intestinais e linhas celulares da origem neyronal (por exemplo, NG108-15). Nas propriedades farmacológicas 5-HT3-retseptory diferenciam-se de outros serotonin receptores; possivelmente, há vários subtipos destes receptores que se diferenciam em várias combinações de subjunits.
Funções mentais. Os meios que produzem receptores serotonin e várias modificações de funções mentais. Muitos modelos experimentais destinados para uma estimativa provisória da estimulação ou bloqueio de atividade da medicina em relação a estes ou aqueles receptores, são baseados em uma pesquisa de tais atos de motivo estereotípicos como, por exemplo, reflexo. As técnicas de Behaviouristic (por exemplo, um método seleto de medicina) permitem sugerir sobre um componente subjetivo de reações a drogas psychotropic. Estas técnicas também usam-se para uma pesquisa da medicina que produz transferência de serotonin, em determinados alucinógenos (veja abaixo). A análise do número enorme dos trabalhos dedicados à influência de serotonin no comportamento está além do nosso livro, e só pararemos naqueles experimentos que têm uma ligação direta com a psicopatologia da pessoa. Para o conhecimento mais detalhado com este sujeito é possível recomendar às revistas perfeitas Glennon e Lucki (1988), Zifa e Fillion (1992), Koeketal. (1992).
Ande de bicicleta um sonho é vigilância. A regulação de um ciclo um sonho é vigilância tornou-se uma das primeiras funções mentais para as quais o papel serotonin se estabeleceu exatamente. Depois do trabalho clássico em gatos que se executa por Mouret e ñîò ð. (Mouret et al., 1967), o jogo de dados que a depleção de estoque serotonin por meio de causas a insônia eliminada entrando de um predecessor de serotonin 5 de aparecido. Também pareceu que tryptophan e não stimulators seletivo de receptores serotonin encurtam o tempo do adormecimento e estendem a duração geral de um sonho. Blockers de receptores serotonin pode subir tanto, e abaixar uma ação de um sono lento profundo que, provavelmente, se causa pela ação em subtipos diferentes de receptores serotonin (Wasquier e Dugovic, 1990). Tanto em animais como em blockers 5-HT2A-and 5-Nt2s-retseptorov (por exemplo, riganserin) um tanto confiantemente causam o aumento em uma ação de um sono lento profundo.
Agressão e impulsividade. Os dados obtidos tanto em animais como na pessoa, demonstre que um serotonin desempenha um papel importante no comportamento agressivo e impulsivo. Em muita comunicação de pesquisas clínicas entre o nível baixo de 5 - do ácido hydroxyindolacetic em SMZh e tal comportamento mostra-se (Brown e Linnoila, 1990). Deste modo, a depressão deste nível ata-se a tentativas impulsivas do suicídio (mas não com pensamentos suicidas; Virkkunen et al., 1995). Bem como em caso de todos outros efeitos de um serotonin, os animais não têm dados finais sobre a influência de um serotonin no comportamento agressivo ainda embora a suposição de tal influência se comprove bastante. Recentemente houve apoio de dados genético e a dilatação de tais visões. Os receptores de Serotonin investigados por métodos da engenharia genética 5-HT) o V-receptor foi o primeiro de. Por uma recombinação homóloga a linha de ratos que têm um gene que codifica este receptor recebeu-se foi inactivated (Saudau et al., 1994). Em tais animais a agressão mais aguda desenvolveu-se o que fala de um papel destes receptores ou em acontecer aos contornos de neuronic que são responsáveis pelo comportamento agressivo ou é imediato na maior parte de tal comportamento. Na pessoa a mutação de ponto do gene que codifica MAO A grava-se; a agressão ao mesmo tempo extrema na combinação com o retardo mental também se observa (Brunner etal., 1993). Como apareceu, as implicações correspondentes estão também disponíveis para ratos com as mutações que resultam na deficiência de MAO A (Casos et al., 1995). Estes dados, certamente, confirmam uma hipótese de um papel de perturbações da transferência de serotoni-nergic no comportamento agressivo.
Preocupação e depressão. Eficiência dos meios que influem serotoninergichesky transferência (por exemplo, nervos inibidores da captura de regresso de serotonin), em caso de frustração perturbadora e depressão evidência de benefício de um papel serotonin em pathogenesis destas condições. Contudo em modelos experimentais clássicos destes a frustração recebe resultados ambíguos o efeito da medicina depende de espécies e raça de um animal e a técnica aplicada. Deste modo, agonist parcial de 5-HT1A-retseptorov buspiron (hl. 19), sendo um tranquilizador eficaz na pessoa, não reduz na preocupação dos animais nas experiências entregues por uma técnica de preferência ou evitação; entretanto esta técnica usou-se em caso do desenvolvimento de tranquilizadores. Ao mesmo tempo o buspiron e outro blockers de 5-HT1A-retseptorov têm o efeito anxiolytic bastante eficaz sobre outros modelos da frustração perturbadora (Barrett e Vanover, 1993). Os trabalhos recentes em ratos com os genes inactivated de 5-NT1L-retseptorov também testemunham para o benefício de um papel destes receptores em pathogenesis da preocupação e, possivelmente, depressões (Parques et al., 1998; Ramboz et al., 1998). De outro lado, stimulators de alguns os receptores serotonin, inclusive 5-T2A--, preocupação de causa de 5-HT2C-and 5-HT3-retseptorov tanto em animais experimentais, como na pessoa. O papel destes receptores e em modelos experimentais de uma depressão supõe-se (por exemplo, o desamparo treinado).
A pessoa de dados diretos sobre um papel serotonin em um pathogenesis de uma depressão ainda tem não bastante. Ao mesmo tempo há fatos clínicos muito brilhantes. Deste modo, em pacientes com uns efeitos de depressão de nervos inibidores do regresso serotonin recompreensão rapidamente eliminam-se nas influências que reduzem serotonin nível em um cérebro. Como tais influências servem, por exemplo, a recepção de parachlorphenylalanine ou as bebidas que não contêm um tryptophan, mas rico com ácidos amino neutrais (Delgado etal., 1990). É interessante que estas influências em si mesmo não causem e não agravem uma depressão. Significa que o nível suficiente de um serotonin em um cérebro é necessário em primeiro lugar para a eficiência de nervos inibidores da sua captura de regresso.
Agentes que influem serotonin nível em tecidos
Nos experimentos enviados na clarificação de um papel fisiológico de um serotonin é possível aplicar blockers de receptores serotonin ou agentes que influem serotonin nível em tecidos. Até há pouco na maioria de trabalhos só as drogas do segundo dos grupos mencionados foram mecanismos usados da ação de blockers os receptores serotonin estudaram-se mal.
No momento da baixa manutenção de um tryptophan em uma dieta serotonin nível em um cérebro diminui, no alto teor de aumentos de tryptophan. Como a reação restritiva da síntese de um serotonin catalisa-se, os nervos inibidores desta enzima causam a queda aguda do nível de um serotonin. Muitas vezes use um nervo inibidor seletivo irrevogável de um triptofangidroksilaza parachlorphenylalanine. Abaixo da influência desta substância serotonin nível consideravelmente e por muito tempo diminui, e a manutenção de catecholamins não se modifica.
Parachloramphetamine e outras benzedrinas halogenated causam a emissão de serotonin de trombócitos e neurônios. Em um cérebro depois que lá vem a depressão longa de reservas de serotonin. As benzedrinas de Halogenated usam-se largamente em trabalhos experimentais. Dois deles fenfluramin e decsfenfluramin aplicaram-se como anorecsant, contudo com relação a relatórios sobre a sua ação tóxica cardio em 1998 eles nos EUA retiraram-se da venda. As consequências do uso destes agentes estão à altura do fim não estudado. Nos neurônios cerebrais serotonin causam expresso e longo (até várias semanas) depressão do nível de um serotonin e o conteúdo da proteína, específica para estes neurônios que uma transportadora de um serotonin e triptofangidroksilaza ao mesmo tempo reduz. Pode demonstrar a ação neurotoxin, contudo os sinais da morte de neurônios abaixo da influência de benzedrinas halogenated não se encontram. Derivados de triptamin com deputados adicionais em linho do Indo um anel (por exemplo, 5.7-digidroksitriptamin; ver o figo. 11.1) indubitavelmente causam a morte os neurônios serotonin. A introdução de um 5.7-digidroksitriptamin um animal adulto leva-se à destruição seletiva por terminações serotonin, contudo os corpos dos neurônios não sofre, e dentro de algum tempo as terminações regeneram. Ao contrário, em animais recém-nascidos tanto as terminações como os corpos de neurônios serotonin e por isso neogenesis não acontece perecem.
Os nervos inibidores do regresso serotonin recompreensão, por exemplo fluoxetine pertencem aos agentes que influem precisamente na transferência de serotonin. O mecanismo da sua ação compõe-se na extensão de efeitos do serotonin alocado no momento da iniciação das terminações nervosas. Se junto com estas drogas para entrar em 5 hydroxytryptophan, então as influências de serotonin agudamente amplificam. Os nervos inibidores do regresso serotonin recompreensão são um dos antidepressivos mais modernos e comuns. O nervo inibidor do regresso serotonin recompreensão, Noradrenalinum e Dofaminum sibutramin aplica-se como anoreksant. Em um organismo dois metabolites ativos que, provavelmente, e têm o efeito terapêutico formam-se dele. Ainda não é claro, influência no que o mediador causou ao efeito de sibutramin.
Os nervos inibidores de MAO e Reserpinum pertencem a não agentes seletivos que influem serotonin nível em tecidos. Os nervos inibidores de MAO bloqueiam o caminho principal de um metabolismo de um serotonin, e Reserpinum causa a sua emissão do armazém de neurônio com o atrito subsequente. Todos estes agentes levam à depressão expressa da manutenção de serotonin, contudo ao mesmo tempo no mesmo grau também o nível de reduções de catecholamine. Por isso, como os reagentes dos nervos inibidores de análise farmacológicos de MAO e Reserpinum usam-se raramente. Aplicaram-se na psiquiatria: Reserpinum como narcoleptic, e nervos inibidores MAO como antidepressivos.

 Carreta
Carreta